
Ossicones are columnar or conical skin-covered bone structures on the heads of giraffe and okapi (Fig 1). Giraffe ossicones consist of a bone core covered with skin and attached to the skull with connective tissue. The bone, skin, and connective tissue of ossicones are all living growing tissues with blood vessels and nerves. All male and female giraffes have a pair of ossicones on the parietal bones of the skull. Males also usually have a single ossicone on the frontal bone. At around the age of sexual maturity, ossicones fuse with the underlying skull bone and growth continues as bone is added to the ossicone surface beneath the skin. Ossicones play a role in giraffe sexual behavior and may also be used for temperature regulation. Male–male combat over access to reproductive females usually consists of giraffes using their head as a club to deliver blows from ossicones to the body and legs of their opponent. The nerve bundles and large blood supply in the ossicones led some researchers to speculate that the structures may play a role in temperature regulation of the brain. I propose the blood vessels and nerves in ossicones could be adaptive traits to promote rapid healing of structures that seem important to male reproductive success. Given the composition of ossicones, and their importance to reproductive behavior, I recommend against any permanent ossicone attachments, and that any methods affecting giraffe ossicones should be carefully reviewed for purpose and needs that are urgent enough to overcome the risk of pain and reproductive harm.

What are Giraffe Ossicones?
Ossicones are columnar or conical skin-covered bone structures on the heads of giraffids (giraffe and okapi; Fig. 1) that are unique among mammals and may be the most basic form of existing mammal headgear (for reviews of mammal headgear structure and evolution see Bubenik and Bubenik 1990, Davis et al. 2011, Nasoori 2020). Giraffe ossicones consist of a highly vascularized and enervated bone core covered with vascularized and enervated skin (Ganey et al. 1990). The base of an ossicone is attached to the skull with vascularized enervated connective tissue (Spinage 1968, Ganey et al. 1990).
All male and female giraffes have a pair of parietal ossicones on the parietal bones of the skull (Lydekker 1904, Spinage 1968). Males also usually have a single median ossicone on the frontal bone that is larger in northern animals and smaller in southern giraffes (Lydekker 1904, Spinage 1968, Seymour 2001). Giraffes can also have additional small paired occipital ossicones on the occipital bones, small paired orbital ossicones above the eyes, and azygous ossicones (Lydekker 1904, Spinage 1968).
Ossicones begin formation in utero as skin-covered connective tissue (Lankester 1907, Ganey et al. 1990). The bony core of ossicones starts forming after birth, beginning as a nodule of bone tissue formed by ossification of dense connective tissue (sesamoid bone; Hall 2015) beneath the skin and distinct from the bone of the skull (Ganey et al. 1990). Ossicones remain detached for a number of years and grow at the skull-ossicone interface and by depositing bone on the ossicone surface beneath the skin (Spinage 1968, Churcher 1990). At around the age of sexual maturity, ossicones fuse with the underlying bone and growth ceases at the skull-ossicone interface, but lamellar deposition on the ossicone surface beneath the skin continues and a knob end may form (Spinage 1968, Churcher 1990).
In males, continuous ossification of the skull and ossicones result in ossicones that are heavier (Mitchell et al. 2013) and larger than in females (Owen 1839, Solounias 2007). Male ossicones have an average circumference of 22 cm and a length of 18 cm (Spinage 1968).
What are the purposes of giraffe ossicones?
There is no clear-cut answer to the question of ossicone purpose. The most prominent theories involve thermoregulation and combat to establish male dominance for reproduction.
Ganey et al. (1990) suggested that the nerve bundles within the horn would be capable of providing a mechanism for sampling the external environment, or preferentially shunting the blood to the external carotid artery if the need for a cooler temperature was sensed at the hypothalmus. Ganey et al. (1990) observed that ossicones are supplied by a surprisingly big branch of the cornual artery, and their venous drainage channels traverse the spongiform bone. The blood supply was described as far more than would be needed to supply the nutrients for the skin covering or the bony core. Thus Ganey et al. (1990) suggested that the ossicones act to dissipate heat to the environment and the veins return cool blood to the venous sinuses in the spongiform bone, providing a layer of cooler temperature between the brain and the environment.
Giraffes are long lived (approximately 25–28 yr; Dagg and Foster 1976) and slow breeding (Bercovitch and Berry 2009). Females reproduce aseasonally throughout the year, with estrous cycling approximately every 15 days, and can become pregnant while still nursing their previous offspring (Dagg and Foster 1976; Bercovitch et al. 2006). Female giraffes in estrous are dispersed over space and time, so reproductive adult males adopt a strategy of roaming among female groups to seek mating opportunities, with periodic hormone-induced rutting behavior approximately every 2 weeks, a temporal scale that would overlap with local cycling females (Pratt and Anderson 1985; Bercovitch et al. 2006; Seeber et al. 2013). Males often fight each other, behavior that is believed to establish dominance hierarchies and access to reproductive females. Male–male combat usually consists of giraffes using their head as a club to deliver blows from ossicones to the body and legs of their opponent (Innis 1958, Coe 1967, Pratt & Anderson 1985).
I propose that an alternative reason for the ample blood and nerve supply of ossicones could be to facilitate the detection and healing of ossicone fractures sustained from combat. The network of blood vessels removes waste and delivers oxygen, nutrients, immune cells, and hormones, and perfusion of the surrounding soft tissues and bone is critical to bone healing (Latta et al 1980, Caruso et al 2000, Ercin et al. 2017). Because head clubbing combat appears to be important to male reproductive success, adaptations that promote rapid healing of these structures could improve the fitness of individual males.
Pain
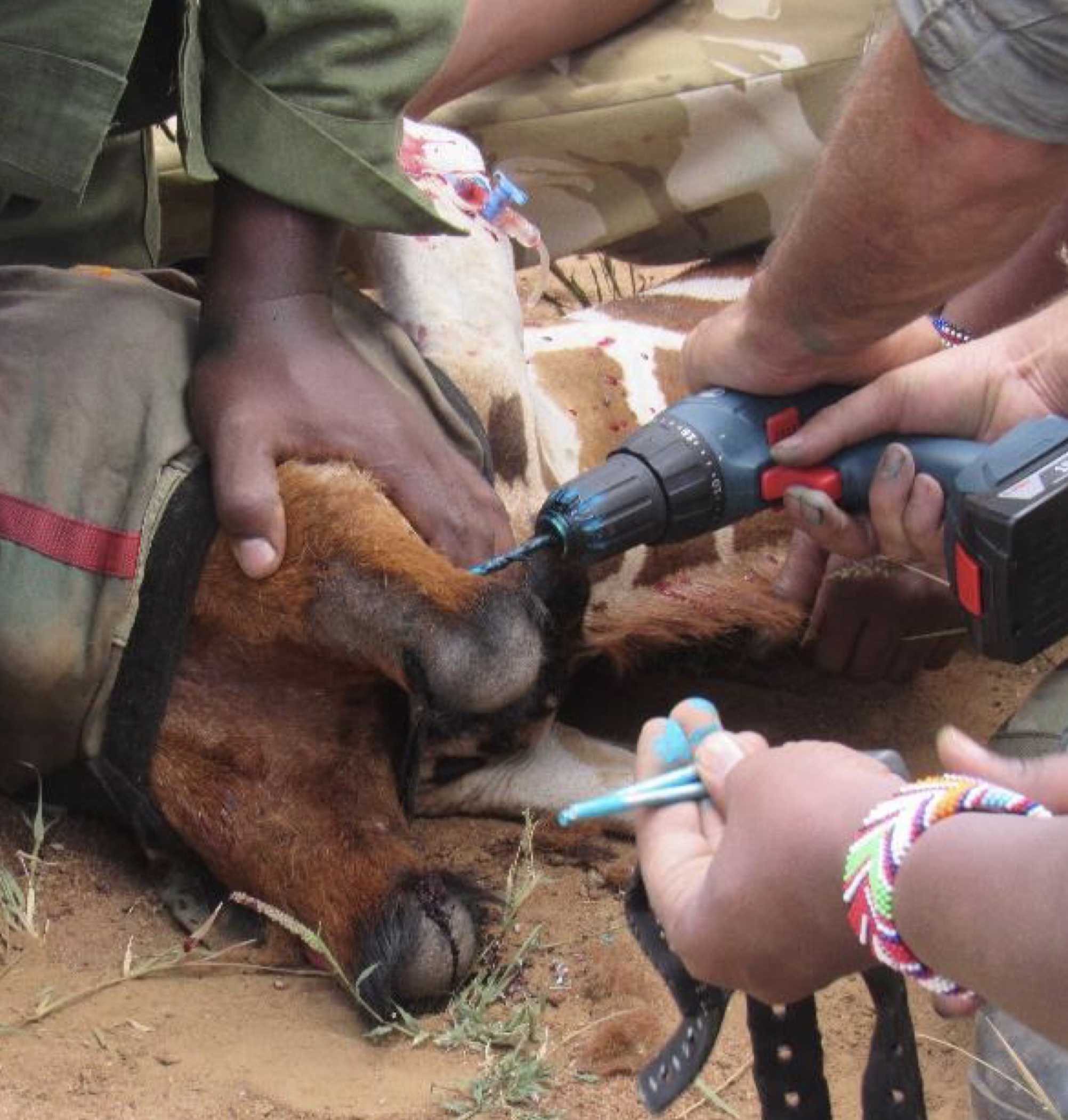
Nociception is an animal’s physical alarm system that protects the body from harm by triggering behavioural and reflex responses to suppress the cause and to limit the negative consequences (Serrie and Serviere 2014). According to Serrie and Serviere (2014), animal pain is defined as an animal’s awareness of an aversive sensory and emotional experience associated with actual or potential tissue damage. The definition applies to vertebrates only and specifies that a painful sensory experience must trigger: protective motor responses (withdrawal of a limb), neurovegetative responses (increased heart rate, higher blood pressure, peripheral vasoconstriction, transitional change in breathing), or learnt avoidance responses (avoidance of a predator or a place associated with an aversive experience, behavioural changes: animal becoming fearful, decreased exploration of a novel place). Giraffes that have had instruments attached to their ossicones using drills and bolts (Fig. 2) demonstrated behavioral changes, perhaps linked to the experience.
Institutional Animal Care and Use Committees (IACUC) provide oversight and ensure appropriate review of the uses of vertebrate animals in teaching, testing, and research. Given the neurological composition of ossicones, and their importance to reproductive behavior, I recommend against permanent instrument attachment to ossicones. Any methods affecting giraffe ossicones should be carefully reviewed for purpose and needs that are urgent enough to overcome the risk of pain and suffering to giraffes. Claims that ossicones are not ennervated, such as those made by Giraffe Conservation Foundation, should be considered inaccurate. Any instrumentation technique concerning ossicones should ensure adequate anesthetic and pain relief is administered to minimize animal discomfort.
Literature cited
Bercovitch FB, Bashaw MJ, del Castillo SM. 2006. Sociosexual behavior, male mating tactics, and the reproductive cycle of giraffe Giraffa camelopardalis. Hormones and Behavior 50:314‒321.
Bercovitch FB, Berry PSM. 2009. Reproductive life history of Thornicroft’s giraffe in Zambia. African Journal of Ecology 48:535‒538.
Bubenik GA, Bubenik AB. 1990. Horns, Pronghorns and Antlers, Evolution, Morphology, and Social Significance. Springer-Verlag, New York, USA.
Caruso G, Lagalla R, Derchi L, Iovane A, Sanfilippo A. 2000. Monitoring of Fracture Calluses with Color Doppler Sonography. Journal of Clinical Ultrasound 28:20–27.
Churcher CS. 1990. Cranial appendages of giraffoidea. In Horns, pronghorns, and antlers: evolution, morphology, physiology, and social significance (eds GA Bubenik & AB Bubenik), pp. 180–194. New York, NY: Springer.
Coe MJ. 1967. ‘‘Necking’’ behaviour in the giraffe. Journal of Zoology 151:313–321.
Dagg AI, Foster JB. 1976. The giraffe: its biology, behavior, and ecology. Van Nostrand Reinhold, New York, USA.
Davis EB, Brakora KA, Lee AH. 2011. Evolution of ruminant headgear: a review. Proceedings of the Royal Society B: Biological Sciences 278:2857–2865.
Ercin E, Hurmeydan OM, Karahan M. 2017. Bone anatomy and the biologic healing process of a fracture. In Bio-orthopaedics (eds A Gobbi, J Espregueira-Mendes, JG Lane, M Karahan) pp. 437–447. Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-662-54181-4_34
Ganey T, Ogden J, Olsen J. 1990. Development of the giraffe horn and its blood supply. The Anatomical Record 227:497–507.
Hall BK. 2015. Bones and Cartilage : Developmental and Evolutionary Skeletal Biology, Elsevier Science & Technology. ProQuest Ebook Central, https://ebookcentral.proquest.com/lib/pensu/detail.action?docID=1910043.
Innis AC. 1958. The behaviour of the giraffe, Giraffa camelopardalis, in the eastern transvaal. Proceedings of the Zoological Society of London 131:245–278.
Lankester ER. 1907. On the origin of the lateral horns of the giraffe in foetal life on the area of the parietal bones. Proceedings of the Zoological Society of London 77:100–115.
Latta LL, Sarmiento A, Tarr RR. 1980. The rationale of functional bracing of fractures. Clinical Orthopedics 146:28.
Lydekker R. 1904. On the subspecies of Giraffa camelopardalis. Proceedings of the Zoological Society of London 74:202–227.
Mitchell G, Roberts D, Van Sittert S, Skinner J. 2013. Growth patterns and masses of the heads and necks of male and female giraffes. Journal of Zoology 290:49–57.
Nasoori A. 2020. Formation, structure, and function of extra-skeletal bones in mammals. Biological Reviews 95: 986–1019. doi: 10.1111/brv.12597
Owen R. 1839. Notes on the anatomy of the Nubian giraffe. The Transactions of the Zoological Society of London 2:217–243.
Pratt DM & Anderson VH. 1985. Giraffe social behaviour. Journal of Natural History 19:771–781.
Seeber PA, Duncan P, Fritz H, Ganswindt A. 2013. Androgen changes and flexible rutting behaviour in male giraffes. Biology Letters 9:20130396.
Serrie A, Servière J. 2014. Pain: definitions, concepts and mechanisms in humans and farm animals. Advances in Animal Biosciences 5:297–309.
Seymour RS. 2001. Patterns of Subspecies Diversity in the Giraffe, Giraffa camelopardalis (L. 1758): Comparison of Systematic Methods and Their Implications for Conservation Policy. PhD thesis, University of Kent at Canterbury.
Solounias N. 2007. Family Giraffodae. In The Evolution of Artiodactyls (eds D Prothero & S Foss), pp. 257–277. JHU Press, Baltimore, USA.
Spinage CA. 1968. Horns and other bony structures of the skull of the giraffe, and their functional significance. East African Wildlife Journal 6:53–61.